صفحه اصلی > روماتولوژی > واکنشهای ازدیاد حساسیت
جدول محتوا
واکنشهای ازدیاد حساسیتی
خلاصه
واکنشهای ازدیاد حساسیتی به پاسخهای سیستم ایمنی غیرطبیعی برمیگردد که به بدن آسیب میرسانند. واکنشهای آلرژی مختلف علیه یک ماده خارجی ایجاد میشود؛ درحالی که بیماریهای خودایمن از پاسخهای ایمنی غیرطبیعی به آنتیژنهای داخلی بدن نشأت میگیرد. واکنش علامتدار فقط در یک فرد حساس شده ایجاد میشود، بنابراین فرد باید قبلاً با آنتیژن مورد نظر مواجه بدون علامت داشته باشد. این واکنشها به 4 گروه تقسیم میشوند: ازدیاد حساسیت تیپ1 به واکنشهای آلرژیک سریع علیه آلرژنهای خارجی (میوهها، گردههای گل، پشم حیوانات) که میتواند به صورت کهیر، آسم و در موارد شدیدتر شوک آنافیلاکسی ظاهر شود. در واکنش ازدیاد حساسیت تیپ 2 (آنمی همولیتیک خودایمن، سندرم گود-پوسچر) سلولهایی که توسط آنتیبادیها احاطه شدهاند، به واسطه فاگوسیتها بلیعده میشوند. واکنش ازدیاد حساسیت تیپ 3 (واسکولیتها، گلومرولونفریت) ناشی از رسوب کمپلکسهای ایمنی آنتیژن-آنتیبادی در بافت مذکور و آسیب به آن بافت است. واکنشهای ازدیاد حساسیت تیپ 4 (تست پوستی توبرکولین، درماتیت) واکنشهای تأخیری وابسته به لنفوسیتهای حساس شدهی T هستند و به آنتیبادیها مرتبط نمیباشند. لازم به ذکر است که در بعضی از حساسیتها مانند حساسیت به ماده کنتراست، در همان برخورد اول ماستسلها هیستامین را آزاد میکنند؛ این واکنشها، واکنشهای ازدیاد حساسیتی غیرآلرژیک نامیده میشوند.
{kind=link}
نگاه کلی
- تعریفها:
- واکنشی که در آن سیستم ایمنی بدن سبب اثر مضری برروی بدن میشود.
- آلرژی: پاسخ ایمنی غیرطبیعی به محرکهای محیطی بیخطر (غذا، گردوغبار، پشم گوسفندان)
- بیماری خودایمن: پاسخ اتوایمیون غیرطبیعی برضد یک آنتیژن که درواقع بخشی از بدن است.
- استیجها:
- حساسیت زایی: تماس بیعلامت ابتدایی با یک آنتیژن
- اثر: پاسخ ایمنی مضر به دنبال حساسیت و پاسخ آنتیژنی به دنبال آن
- تایپها: واکنشهای ازدیاد حساسیتی به چهارگروه تقسیم میشوند.
| خلاصه پاتوفیزیولوژی | مثالها | |
| تیپ I: فوری💬 |
|
|
| تیپ II: سیتوتوکسیک |
|
|
| تیپ III: کمپلکسهای ایمنی |
|
|
| تیپ IV: تأخیری (ناشی از فعالیت T-cell) |
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
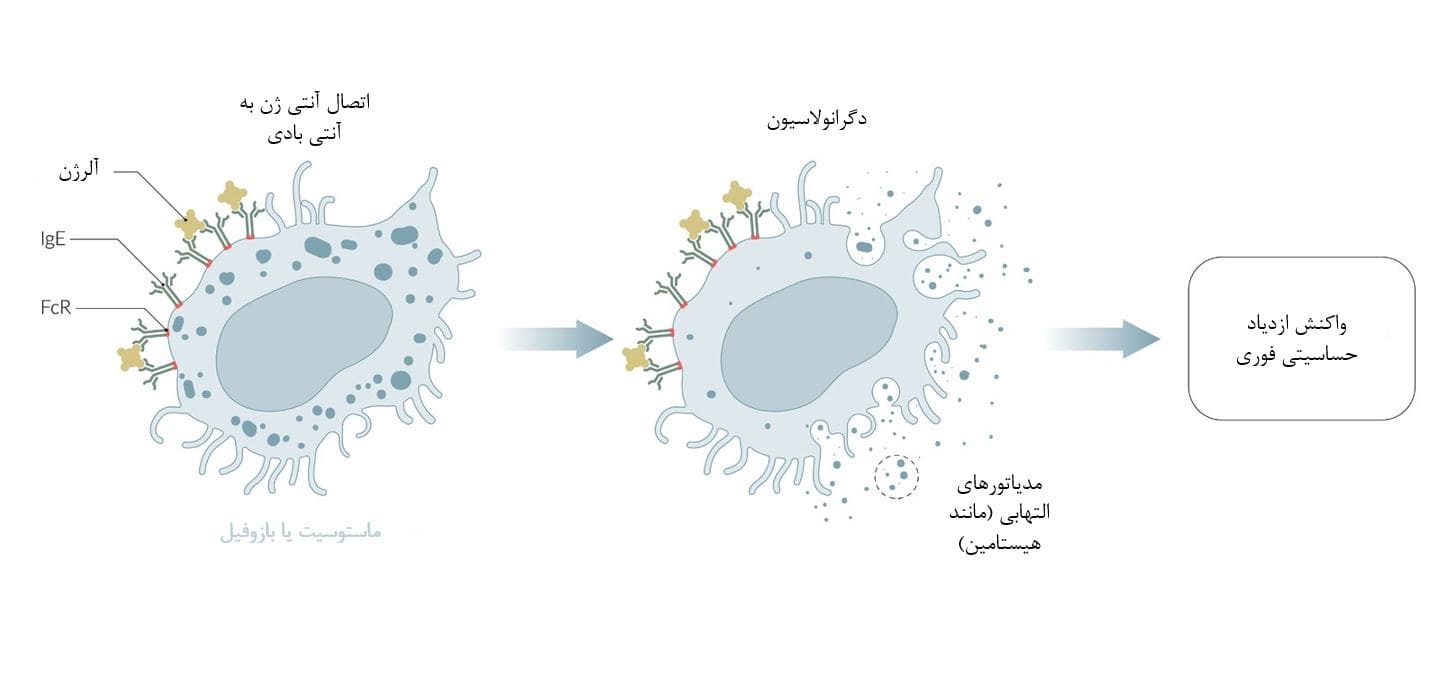
IgE معمولاً به گیرندههای Fc یا FcR در سطح سلولهای ایمنی مانند ماستوسیتها و بازوفیلها متصل میشود. اتصال متقابل یک ماده حساسیت زا با IgE به سلولهای ایمنی سیگنال میدهد تا دگرانوله شوند و واسطههای التهابی (مانند هیستامین) آزاد کنند که بلافاصله واکنش ازدیاد حساسیت را ایجاد میکنند.
این پاتوفیزیولوژی واکنشهای ازدیاد حساسیت نوع I است (مانند آنافیلاکسی، آلرژی غذایی، واکنشهای دارویی)، که میتواند به قدری شدید باشد که منجر به شوک آنافیلاکتیک شود.
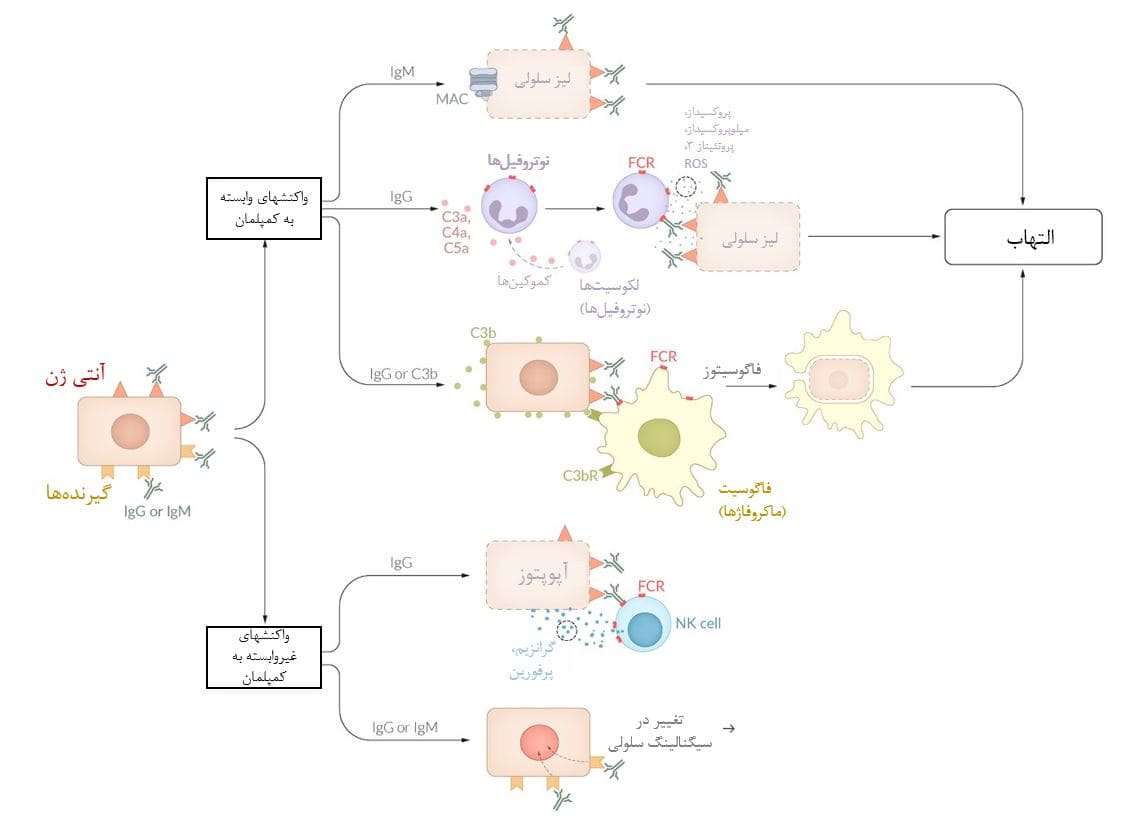
IgM یا IgG آنتیژنهای خاص سطح سلول را تشخیص داده و به آنها متصل می شود. این اتصال متقابل آنتی ژن و آنتی ژن آبشار مکمل را در نتیجه التهاب و واکنش حساسیت نوع II فعال می کند (به عنوان مثال، واکنش انتقال همولیتیک حاد، کم خونی همولیتیک خودایمن)
واکنشهای وابسته به کمپلمان:
– IgM متصل به آنتی ژن یک فعال کننده قوی کمپلمان است و منجر به تولید کمپلکس حمله غشایی (MAC)، محصول نهایی آبشار کمپلمان میشود. MAC سوراخهایی را در غشای سلول ایجاد میکند و باعث سیتولیز سلول حامل آنتی ژن میشود.
– IgG متصل به آنتی ژن به گیرندههای Fc یا FcR در سطح نوتروفیلها و ماکروفاژها متصل میشود و به ترتیب باعث تحریک فعالیت سیتولیزی و فاگوسیتوزی آنها میشوند. IgG متصل به آنتی ژن همچنین آبشار کمپلمان را فعال میکند که چندین پروتئین مکمل فعال تولید میکند؛ از جمله C3b که به غشای سلول متصل به آنتی ژن متصل میشود و آن را برچسب گذاری میکند تا توسط ماکروفاژها فاگوسیته شود (یعنی opsonization).
واکنشهای مستقل از کمپلمان:
– IgG همچنین در واکنشهای مستقل از کمپلمان، مانند آپوپتوز نقش دارد و همراه با IgM سیگنالینگ سلولی را تغییر میدهد که منجر به اختلال در عملکرد سلولی و مرگ آن میشود.
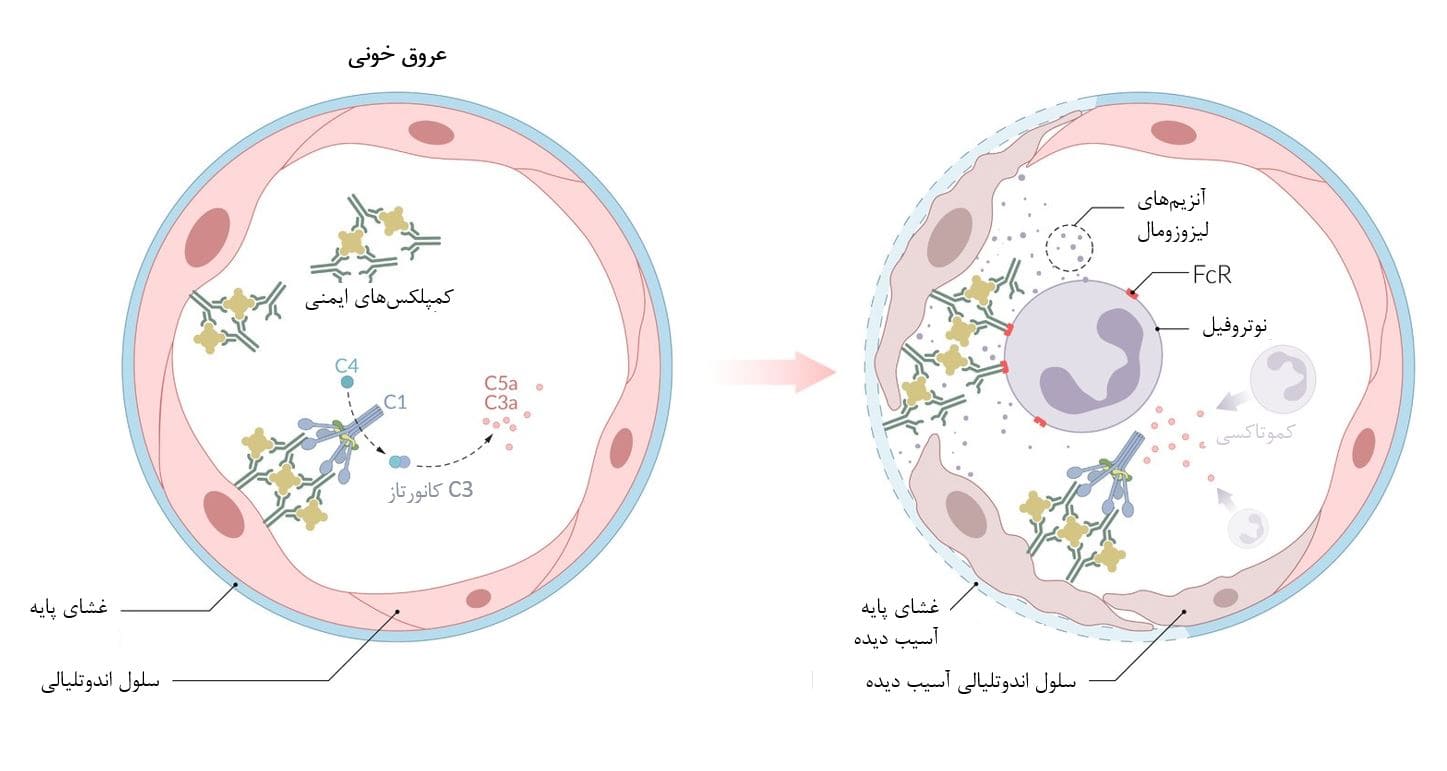
کمپلکسهای ایمنی (کمپلکسهای آنتیژن-آنتیبادی) در گردش خون تشکیل میشوند و در بافتها ترجیحاً درون اندوتلیوم عروقی، اندامهای خاصی (مانند کلیه، ریه) و غشاهای سروزی (مانند سینوویوم، پلور، پریکارد) رسوب میکنند.
کمپلکسهای ایمنی رسوب یافته، آبشار کملپمان را فعال میکنند و در نتیجه باعث کموتاکسی (جذب نوتروفیلها) میشوند. IgG متصل به آنتی ژن به گیرندههای Fc یا FcR در سطح نوتروفیلها متصل میشود و سپس آنزیمهای لیزوزومی آزاد میکند، که به بافتهای موضعی آسیب میرساند.
محل رسوب کمپلکس ایمنی، ویژگیهای بالینی یک واکنش حساسیت نوع III را تعیین میکند (مانند واکنش آرتوس، واسکولیت، گلومرولونفریت، سینوویت، پنومونیت).
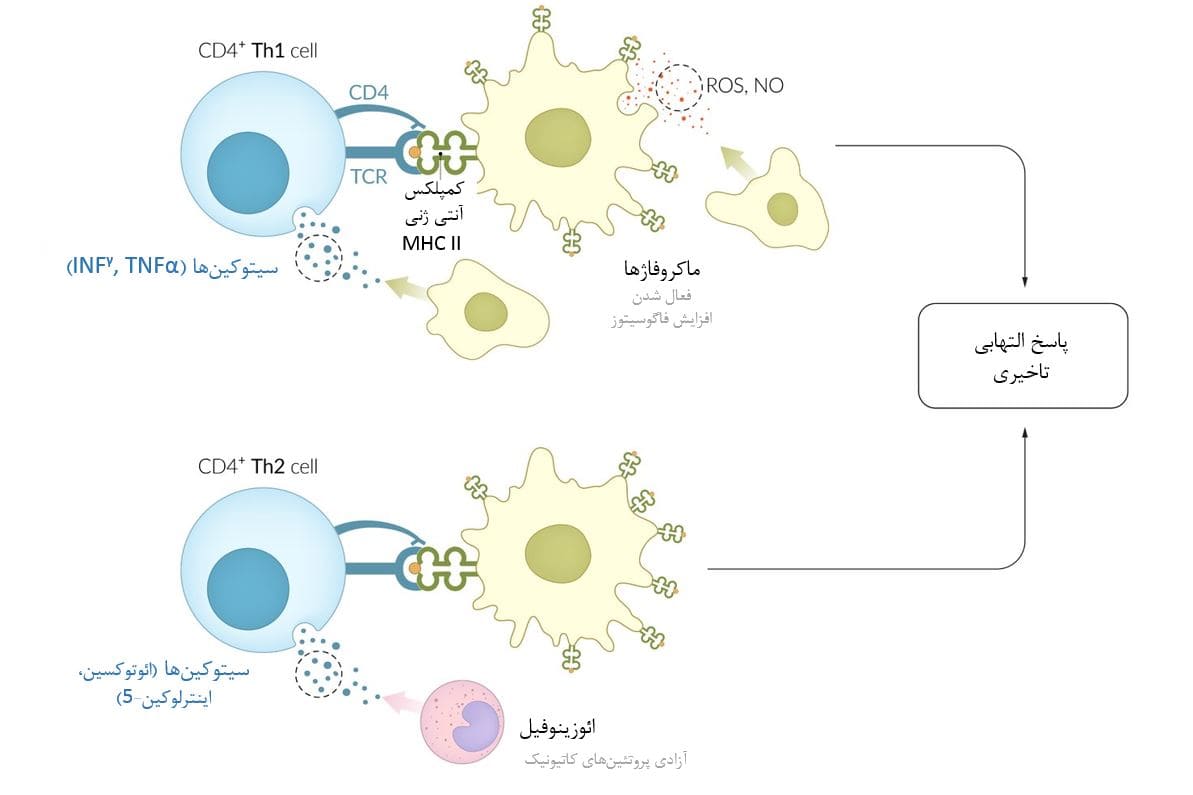
واکنشهای حساسیت نوع IV برخلاف همه واکنشهای حساسیت دیگر که بواسطه آنتی بادی هستند، توسط سلولهای T واسطهگری میشوند. سلولهای کمکی T (سلولهای +CD4؛ سلولهای Th) عمدتاً با جذب سلولهای التهابی با واسطه سیتوکین در محل سلولهای هدف عمل میکنند.
گیرندههای سلول T یا TCR موجود در سلولهای CD4 + حساس شده، آنتیژنهای خاص اگزوژن (مانند باکتریها، ویروسها) را تشخیص داده و به آنها متصل میشوند. این آنتیژنها به MHC II (یک پروتئین غشایی موجود در سطح سلولهای APC یا ارائه دهنده آنتی ژن)، مانند ماکروفاژها متصل هستند. سلولهای Th بر اساس سیتوکینهایی که تولید میکنند به دو زیر مجموعه زیر تقسیم میشوند:
– عفونتهای باکتریایی و ویروسی عمدتاً سلولهای Th1 را فعال میکنند که سیتوکینهایی را آزد میکنند که نوتروفیلها و ماکروفاژها را تحریک میکنند (به عنوان مثال IFNγ ،TNFα). ماکروفاژهای فعال گونههای واکنشگر اکسیژن (ROS) و اکسید نیتریک (NO) را آزاد میکنند، که به تخریب عوامل بیماری زا و فاگوسیتوز آنها کمک میکند.
– عفونتهای کرمی یا انگلی غالباً سلولهای Th2 را فعال میکنند که سیتوکینهای فعال کننده ائوزینوفیلها را آزاد میکنند (مثلاً IL5). ائوزینوفیلهای فعال شده پروتئین اساسی اساسی (MBP) آزاد میکنند که به تخریب کرمها کمک میکند و به سلولهای ماستوسیت سیگنال میدهد تا هیستامین آزاد کنند.
سیتوکینها، ROS و MBP سلولهای التهابی بیشتری را به محل واکنش جذب میکنند و پاسخ التهابی را تقویت میکنند، که معمولاً حدود 48 ساعت پس از قرار گرفتن در معرض آنتی ژن (یعنی ازدیاد حساسیت تأخیری) آشکار میشود.
واکنشهای حساسیت نوع IV برخلاف همه واکنشهای حساسیت دیگر که بواسطه آنتی بادی هستند، توسط سلولهای T واسطهگری میشوند. سلولهای T سیتوتوکسیک (سلول +CD8) عمدتاً با القای آپوپتوز سلول هدف عمل میکنند.
گیرندههای سلول T یا TCR واقع در سلولهای +T CD8 حساس شده، آنتیژنهای سنتز شده اندوژن (معمولاً آنتیژنهای ویروسی) را شناسایی میکنند و به MHC I (پروتئین غشایی موجود در سطح سلولهای هسته دار و پلاکتها) متصل میشوند. اتصال سلولهای +T CD8 به آنتیژن باعث تحریک دو مسیر سیگنالینگ میشود. این مسیرها با فعال سازی آنزیمهای داخل سلول کاسپاز و از بین بردن عوامل بیماری زای داخل سلولی منجر به آپوپتوز سلول آلوده میشوند.
– مسیر ذاتی (مسیر اصلی): سلولهای CD8 + T گرانولهای پروتئاز (گرانزیم، پرفورین) را در سلول آلوده آزاد کرده و باعث آپوپتوز می شوند.
– مسیر خارجی: لیگاند Fas یا FASL موجود در سلولهای +T CD8 به گیرندههای Fas سلولهای هدف متصل شده و باعث آپوپتوز میشود.
مسیر سیتوکین: سلولهای T فعال شده +CD8 همچنین سیتوکینهایی را آزاد میکنند که تکثیر ویروس را کاهش میدهد و بیان MHC I را افزایش میدهد تا تشخیص آنتی ژنها تقویت شود. سیتوکینها همچنین ماکروفاژها را به محل واکنش جذب میکنند و باعث ایجاد واکنش التهابی میشوند که معمولاً حدود 48 ساعت پس از قرار گرفتن در معرض آنتی ژن (یعنی ازدیاد حساسیت تأخیری) آشکار میشود.